发布时间:2019-07-03
喧闹酒会,若你凝视交谈对象,亦能听清对方的轻言细语;而即便宁静夏夜,如果你目光游离,心不在焉,耳边的话语也会难以察觉。一盘色泽艳丽同时香气扑鼻的菜肴,顿时让食欲大开;但若有香无色,滋味又会大打折扣。清醒的时时刻刻,每个人都在接受着多种感觉信息输入,我们用眼、耳、鼻、舌等器官分别获取着视、听、嗅、味等信号的输入。这些由单个感觉通道接受和加工感觉信号的形式被称为单模态感觉处理。然而,我们感知的绝大部分外部事件,同时会具有多个模态的感觉信息。不难想象,为了使对环境事件的感知更加准确,大脑势必将这些被分解到单模态的信息,以某种方式重新合拢加工,形成相对于单模态信息更加有效的感知。多感觉信息合拢加工的过程,是大脑的一个基本功能。
虽然是大脑的基本功能,目前国际上对多感觉信息加工的研究多集中在行为学与心理学层面,对其神经环路机制的研究还少有涉足,还没有发现一个相对细致的神经机制来解释任何一种多感觉信息加工的发生过程。两个原因造成了这种研究现状。其一是多感觉信息加工发生的普遍性,它时刻在我们大脑中进行,在行为学、心理学层面只需细致地观察检测,就能发现有趣且重要的现象。例如1954年发现的“鸡尾酒会效应”,Sumby 和Pollack 从心理物理学角度估测视觉输入可等价于15~20 分贝的听觉输入。又如McGurk 和MacDonald 在1976 年报道的“McGurk effect”中,描述了看到的发音口型与真实声音不一致时,会产生错觉,“听”到介乎二者之间的发音。第二个原因,则是介导多感觉信息加工的神经环路异常复杂,这直接造成了对多感觉信息加工的研究难以深入到神经机制层面的困境。我们知道神经系统是与宇宙星系等量齐观的复杂系统,即便在小鼠这样的小型哺乳动物中,介导单个感觉模态的神经环路已经十分复杂,从外周往中枢,也只是在最初的几个节点相对清楚。如果要涉及到多感觉模态的加工,在结构上必然涉及到单模态感觉通路的相互汇集,其中的复杂度更是不言而喻。
尽管如此,随着神经科学的发展,多感觉信息加工神经机制的研究得到了越来越多科学工作者的重视,并且逐步对其有了更多的了解。根据已有的研究工作,可以宏观上将多感觉信息加工分为两个大类。一是多感觉信息整合,即两个感觉通道的神经环路汇集到同一个下游目标节点上,从而将各自的神经电信号贡献到下游节点的神经活动中,形成一种新的合并的感知。另一种多感觉信息加工的模式是跨感觉模态调节,即一个模态的感觉输入,会通过改变第二个感觉模态通路的工作状态来调节该感觉通路的信号加工。
在多感觉信息整合的研究中,从20 世纪80 年代开始,Stein 和他的同事们在猫的上丘中做了开创性的工作。他们通过细胞外记录发现,在猫的上丘中,有些神经元会同时对多个模态的感觉刺激都产生反应,例如视觉刺激和听觉刺激都分别能引起同一个神经元的放电。继而,他们以此为研究对象,分析了当动物同时接收多个感觉模态刺激时,这些上丘神经元如何产生与单个模态刺激不同的反应[1]。接下来,Angelaki 实验室[2] 以及Logothetis 实验室[3]在恒河猴上开展了一系列工作,从多感觉整合的角度,分别在视觉通路和听觉通路中显示了这种多感觉整合现象的存在。而对于跨感觉模态调节的研究,探索工作则很少,在我们工作发表之前,最前沿的研究来自于Schroeder 实验室,他们同样通过监测灵长类皮层中的胞外电活动,发现一个通路的感觉输入虽然不引起其他感觉通路神经元的放电,但可以显著调节这些神经元对其特定感觉输入的反应[4]。上述这些最近的研究进展虽然在行为学研究上往前推进了一步,但由于所涉及神经环路的复杂性,下述三个问题依旧没有答案:(1) 在结构上,多个模态的感觉信息如何由不同的神经环路汇聚到一起;(2) 在机制上,当多个感觉通路的输入汇聚时,经过了怎样的变化,从而实现神经功能的改变;(3) 在动物行为上,这种汇聚形成的神经信号,如何决定或改变动物的行为。
在神经科学乃至整个生命科学的发展史上,新的模式生物的应用往往能带给一个领域克服过往瓶颈的契机,正如果蝇之于神经遗传学领域的开创,正如线虫之于细胞凋亡原理的发掘。斑马鱼,因其所拥有的研究特点,在神经科学功能性研究中的作用日趋重要。首先,斑马鱼拥有相对简单的神经环路。人类大脑中的大部分脑区( 尤其那些功能重要且保守的脑区),都能在斑马鱼的大脑中找到对应的同源脑区。在拥有足够复杂度的同时,斑马鱼幼鱼的大脑神经元总数只有约10 万个,相对于人类大脑拥有近1000 亿个神经元而言,其神经环路相对简单了多个数量级。第二,斑马鱼幼鱼脑部透明,利于在整体动物上进行神经环路功能和形态研究。第三,遗传背景清楚,基因操作工具在斑马鱼中的应用已日渐丰富。结合这三点优势,我们就拥有了回答前面提出的三个问题的可能性:简化的神经环路结构以及遗传操作带来的可视化( 例如对特定感觉通路神经元的荧光标记),会帮助我们找寻多感觉信息汇聚的结构基础;脑部透明和简化的环路,使得我们可以用在体电生理和成像技术,详尽展示目标脑区神经元的工作机理;简化的环路,使得感觉输入到运动输出的神经通路环节较少,易于解析多感觉整合产生的神经功能的改变如何影响动物的行为。
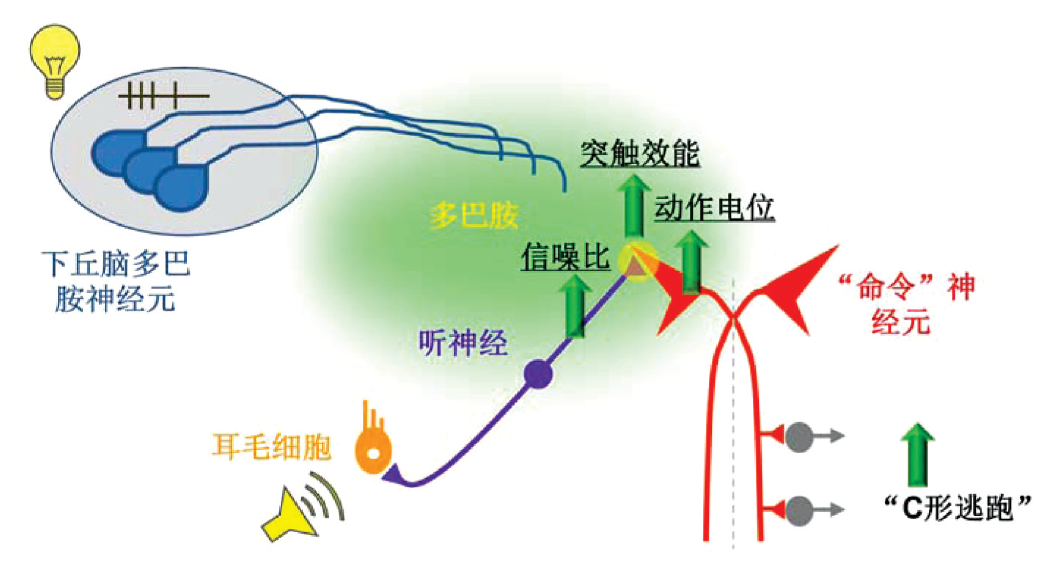
基于这些考虑,我们选用了斑马鱼幼鱼来破解多感觉整合的神经环路机制。首先,我们建立一个稳定表征多感觉整合发生的行为学范式。斑马鱼以及大部分的硬骨鱼中,都存在一种听觉引起的逃跑行为,称之为“C 形逃跑”[5]。由于声音在水中传递速度快、衰减慢,因此相对于其它感觉,听觉在快速判别环境危险过程中更加重要。而“C 形逃跑”便是鱼类远离危险源( 往往伴随突发的声音刺激)的最快行为反应。斑马鱼在接收到威胁性的声音刺激之后的短短20~30 ms 内,其躯体一侧肌肉就会发生大规模的强力收缩,使得其整个躯体尤其头部,朝向刺激源相反的方向转动,继而快速逃离。在此过程中,鱼的身体会弯曲为一个“C”形,由此得名“C 形逃跑”。作为一种事关生死的行为,“C 形逃跑”虽然主要由声音刺激引发,但我们有足够的理由去猜想,它的发生会受到动物所处环境中其他感觉信号( 例如视觉) 的影响。于是,我们通过摸索,建立了一套稳定的发生跨感觉模态调节的行为学范式。在这个行为范式中,闪光视觉刺激本身并不引起任何“C 形逃跑”,但是能大幅度提高随后声音
刺激引起的“C 形逃跑”发生的概率。在此基础上,利用活体电生理记录的方法,我们证实了在执行“C形逃跑”最关键的“命令”神经元上[5],听觉产生的电反应有效地被闪光刺激所调节;对应的,闪光刺激本身并不能在该命令神经元上引起明显的电反应[6]。
基于在动物行为和神经元活动上观察到的跨感觉模态调节现象,接下来我们着手从突触和神经环路两个层次上探究这种调节现象背后的神经机制。首先,运用多种研究手段,包括电生理、药理学、基因操控、荧光追踪等方法,我们发现视觉输入导致在命令神经元上听觉反应的增大源于两步协同的调控。在命令神经元的上一级,即听神经上,与背景噪音相关的自发放电活动被视觉刺激大幅度抑制,而听觉刺激引起的听神经放电则基本不变,从而导致听觉神经信号的信噪比提高;同时,从听神经到命令神经元的突触传递也被视觉刺激所增强。因此,通过这两步协同调节,神经系统实现了在不放大噪音的情况下,选择性地将听觉反应增强,从而提高听觉引发逃跑行为的灵敏度和准确度( 图1)。
那么,在神经环路层次上,视觉输入又是如何作用到听觉运动通路上的呢?其结构基础是什么?通过进一步的实验,我们得到了更加有趣的发现。在脑中,有多种调质系统,它们的细胞体集中,但轴突投射遍布大脑的各个脑区,因此能广泛地调节大脑的功能状态[7]。多巴胺能系统便是调质系统中的一个主要成员。我们发现,视觉引起的上述两步调节都依赖于多巴胺I 型受体的激活。而引起这些受体激活的多巴胺,则来自下丘脑的一个多巴胺能神经元核团。更进一步地,我们证实了这团多巴胺能神经元的确能够被视觉输入激活,从而起到一个视觉输入进入听觉运动通路中继站的作用。由于调质系统广泛地渗入到各种感觉通路,而调质系统又可能接收各种感觉模态的输入,由此我们发现的这种“感觉模态A→ 调制系统→ 感觉模态B”的模式,很有可能是一种普适的神经构架,来广泛介导跨感觉模态的调节。同时,我们发现的先“降噪”再“增效”的两步调节,作为调质系统的作用机制,可以有效地调节神经环路对于感觉信息反应的准确度与灵敏度。这种协同机制是否广泛存在是一个有趣的问题,可以在更广泛的神经环路内予以探究。
这种基于整体动物的研究,赋予了我们在更宏观尺度上进行思考的空间。我们发现的跨模态调节,除了能够增加听觉引起“C 形逃跑”本身,还有什么生态学上的意义呢?结合斑马鱼在自然环境中的生活习性,这种跨模态调节可能具有多方面的生态学意义[8]。比如,斑马鱼常常生活在水草茂密的环境中,这种自然环境除了能提供充足的食物,还能作为掩蔽所为其提供躲避敌害的荫庇。一旦有大型的捕食者进入,势必扰动水草,打破原有静谧。此时快速表征这种入侵的信号,就是视觉上光亮的突然变化。当动物察觉到环境光强的变化时,迅速提高警觉,增强对危险信号感知的灵敏度,从一种休闲的觅食状态,转到另一种高度紧张的备战状态,这是一种很利于生存的策略。更有意思的是,斑马鱼是一种群居动物,拥有极强的鱼群组织。鱼群中,当任何一条鱼,从光线较暗的内部切换巡游到光线较亮的外部时,首先感知到的,就是光强的变化。倘若发生这种光强的提高,鱼儿将提高警觉,鱼群就会形成一个非常有趣的群体组织:内部放松,只需要追随;外部灵敏,充当整个鱼群的“耳目”。
从分子到突触到环路,从环路到行为,再从行为到生态意义的思索,当我们把这整个过程在实验和思路上贯通时,所感受到的美妙沁人心脾,其间一个个小故事更是如星辰,点缀于这条主干周围。最后,简单描述一下这些星辰中的第一个,以飨读者。那是我们探索的起点,其实源于一次偶然的干扰:在尚未开始这个课题的一个下午,当我在记录听觉反应时,一位同事在实验台边的进出,导致了听觉反应的陡然变化。这个变化冲击了我那一时刻的平静,反复模拟同事的进出,思索,并最终确定是视觉的扰动带来这种听觉反应的变化。从那一刻,我们扬帆,开启这一科学问题探索的航程。
苏子赋云:“耳得之而为声,目遇之而成色,取之无禁,用之不竭。”在这一次科研的远航中,我们将苏子之词句做了两层意义的引申。第一层是“目遇”亦可以“为声”,即视觉输入可以调节听觉感知;再深一层,“目遇之为声,取之无禁,用之不竭”则似乎概括了我们发现的历程。“目遇之为声”,从与预想不同的角度去思考,不拘于既有的研究方向,才有了“用之不竭”的思路与探索空间。故以此句与读者共勉:科研路上,用心看,用耳听,目遇亦可为声,无拘无束,用之不竭。
致谢:感谢杜久林研究员对研究工作的指导和对本文的修改。
(发表于《生命科学》201,24:1380-1382)
(核稿:杜久林、李澄宇)
参考文献
Stein BE, Stanford TR. Multisensory integration: current issues from the perspective of the single neuron. Nat Rev Neurosci, 2008, 9: 255-66
Angelaki DE, Gu Y, DeAngelis GC. Multisensory integration: psychophysics, neurophysiology, and computation. Curr Opin Neurobiol, 2009, 19: 452-8
Kayser C, Logothetis NK, Panzeri S. Visual enhancement of the information representation in auditory cortex. CurrBiol, 2010, 20: 19-24
Ghazanfar AA, Chandrasekaran CF. Paving the way forward: integrating the senses through phase-resetting of cortical oscillations. Neuron, 2007, 53: 162-4
Eaton RC, Lee RK, Foreman MB. The mauthner cell and other identified neurons of the brainstem escape network of fish. Prog Neurobiol, 2001, 63: 467-85
Mu Y, Li XQ, Zhang B, et al. Visual input modulates audiomotor function via hypothalamic dopaminergic neurons through a cooperative mechanism. Neuron, 2012, 75: 688-99
Berger B, Gaspar P, Verney C. Dopaminergic innervation of the cerebral cortex: unexpected differences between rodents and primates. Trends Neurosci, 1991, 14: 21-7
Engeszer RE, Patterson LB, Rao AA, et al. Zebrafish in the wild: a review of natural history and new notes from the field. Zebrafish, 2007, 4: 21-40
 附件下载:
附件下载: 